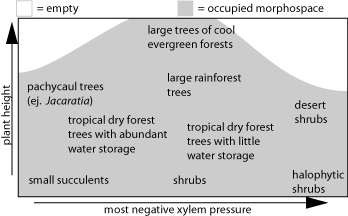
Because of their simple structure and because they can’t run away, we like to work on clades of woody plants of extraordinary morphological diversity. These systems include Moringa (Moringaceae), the simaruba clade of Bursera (Burseraceae), the Mexican clade of Manihot (Euphorbiaceae), and the Beaucarnea- Calibanus clade of Nolinaceae. We are also taking a complementary approach that examines the ways that entire communities fill morphological and functional space. For example, the plants of the Pedregal de San Ángel, which I can see from my office here in Mexico City, span a range greater than that seen by most anatomists in a lifetime: Sedum oxypetalum, whose wood is composed only of vessels and parenchyma, grows adjacent to Dodonaea viscosa, which has extremely reinforced wood that withstands extremely negative xylem pressures. How do the species of a community partition morphological and functional space? Does each species occupy a distinct region? Can guilds of species of similar structure and function be distinguished? We are currently generating phylogenetic, morphological, and functional data for this and other communities to examine these questions. We are also interested in conceptual aspects of evolutionary biology that affect our work. Current projects include understanding the differences between the ways that botanists and zoologists use the term homology, the problem of multiple explanations for the same observation in studies of adaptation, and a review of the conceptual foundations of comparative wood anatomy. We also spend a lot of time fretting about the meaning of branch lengths in phylogenetic comparative methods. Finally, as world petroleum production drops, it is clear that we need to reduce our energy consumption, and meet our needs for food, water, and health care locally. We are involved in exciting projects in rural communities to build a sustainable future together. See also: Our Publications page Courses page |
        |
home - people - research - images of the dry tropics - exploration - acknowledgements
Instituto
de
Biología, Universidad Nacional Autónoma de México
Tercer Circuito s/n, Ciudad Universitaria
Copilco, Coyoacán A. P. 70-367
C. P. 04510, México, D. F.
MÉXICO
(52)
5622-9124
fon (52) 55 5555-1760 fax
molson@ib.unam.mx
